A detailed look at DMRs that hold true across oysters exposed to heat shock.
Updated July 6, 2015 – added three more DMRs (at bottom of post)
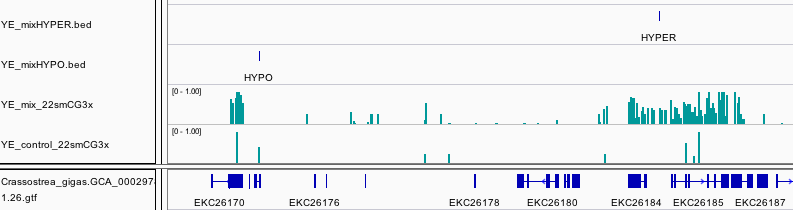
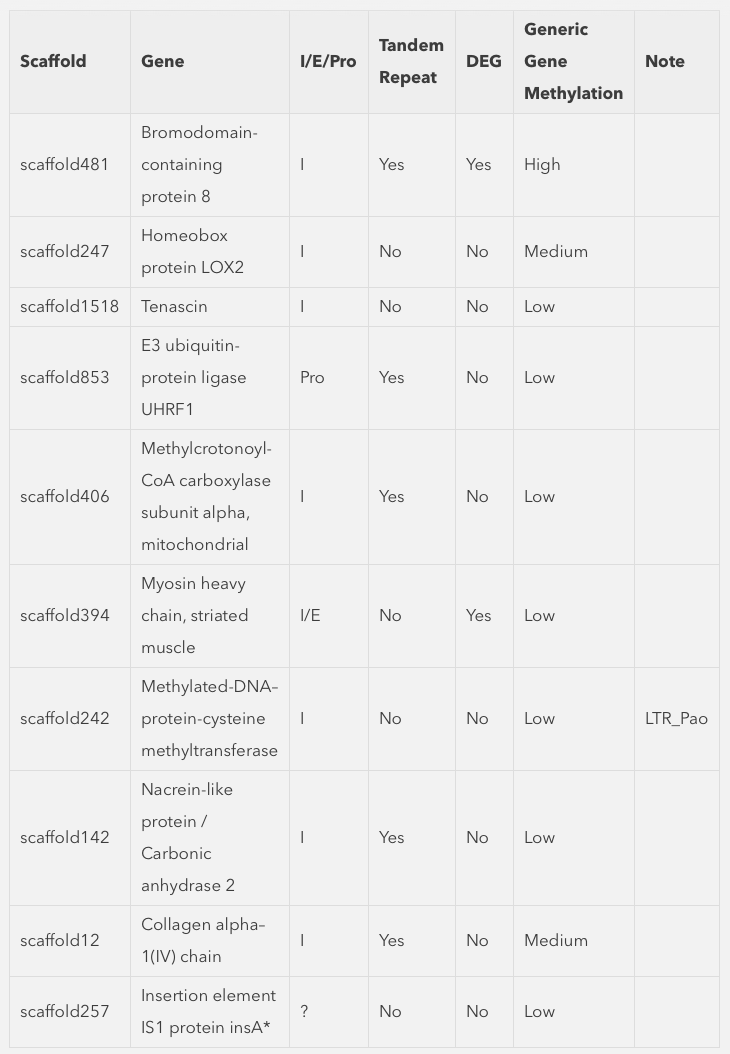
Going down the list, scaffold418_576986 is a feature that overlaps gene EKC36328, Bromodomain-containing protein 8. Specifically the location is in the intron between exon 18 and 19 (total of 20 exons). This gene is differentially expressed, that is expressed at an elevated level following heat shock.
“The precise function of the domain is unclear, but it may be involved in protein-protein interactions and may play a role in assembly or activity of multi-component complexes involved in transcriptional activation [PMID: 7580139].”
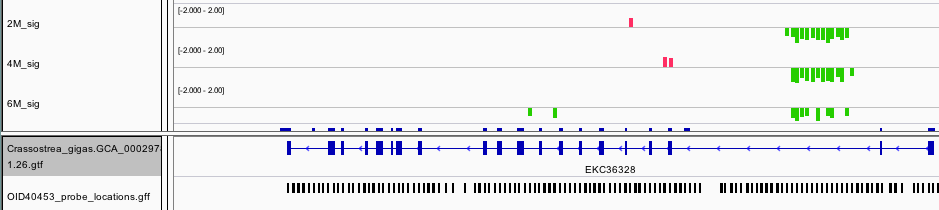
Another DMR that is consistent across oysters is located within the intron of Homeobox protein LOX2. Homeobox are transcription factors often associated with developmental processes.
Significant hypomethylation is also present within the intron of Tenascin, a glycoprotein expressed in the extracellular matrix during stress.
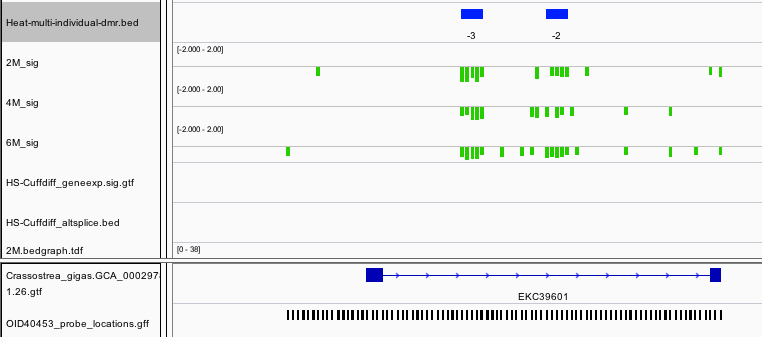
We also found a DMR upstream of E3 ubiquitin-protein ligase UHRF1. Interestingly this is a protein that bridges DNA methylation and chromatin modification.
“Specifically recognizes and binds hemimethylated DNA at replication forks via its YDG domain and recruits DNMT1 methyltransferase to ensure faithful propagation of the DNA methylation patterns through DNA replication. In addition to its role in maintenance of DNA methylation, also plays a key role in chromatin modification: through its tudor-like regions and PHD-type zinc fingers, specifically recognizes and binds histone H3 trimethylated at ‘Lys-9′ (H3K9me3) and unmethylated at ‘Arg-2′ (H3R2me0), respectively, and recruits chromatin proteins. Enriched in pericentric heterochromatin where it recruits different chromatin modifiers required for this chromatin replication. Also localizes to euchromatic regions where it negatively regulates transcription possibly by impacting DNA methylation and histone modifications. Has E3 ubiquitin-protein ligase activity by mediating the ubiquitination of target proteins such as histone H3 and PML. It is still unclear how E3 ubiquitin-protein ligase activity is related to its role in chromatin in vivo.” ~http://www.uniprot.org/uniprot/Q96T88
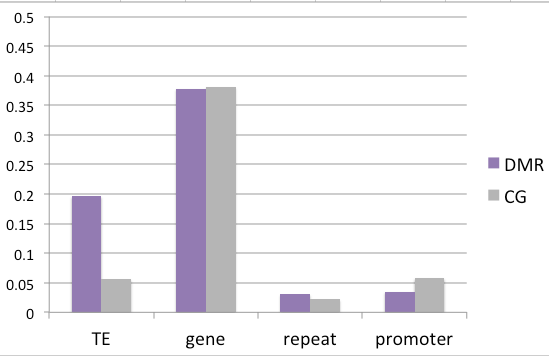
While not classified as a differentially expressed gene, there does appear to be a trend towards increased expression upon heat stress. This occurrence would follow the traditional model where decreased methylation in the promoter region is associated with increased expression.
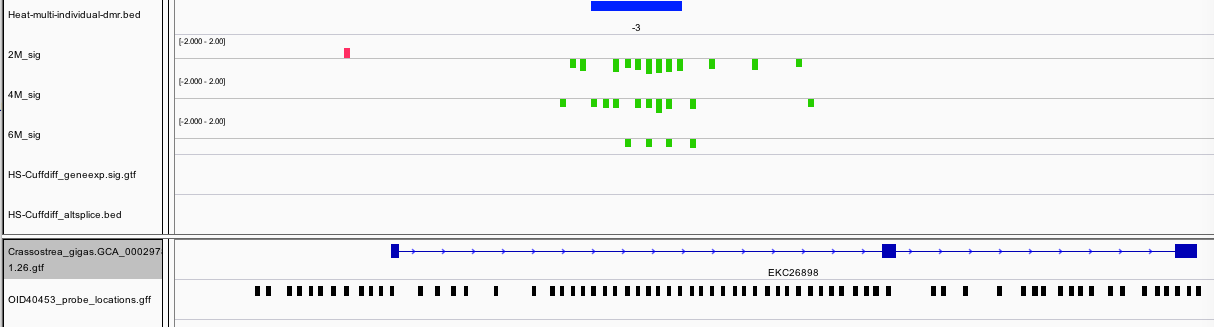
There were three features identified that are in fact within an intron of Methylcrotonoyl-CoA carboxylase subunit alpha, mitochondrial, and enzyme involved in leucine and isovaleric acid catabolism.
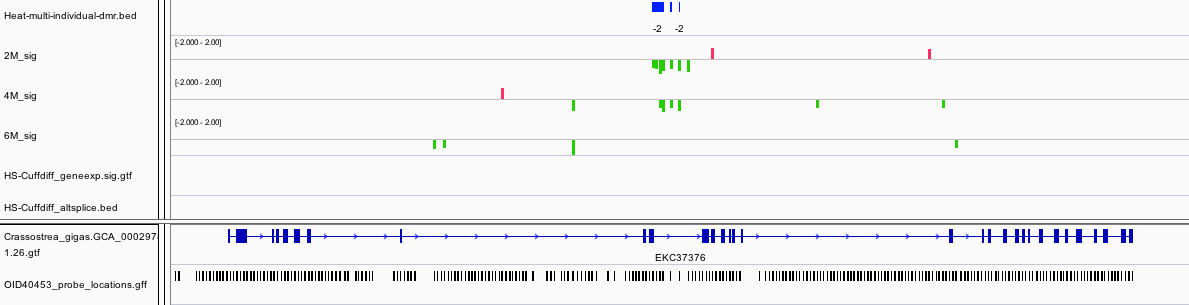
In an another example of a DMR associated with a differentially expressed gene, a DMR that span an intron and exon within Myosin heavy chain, striated muscle. In this case the gene is expressed at a lower level upon heat stress. It is also worth pointing out this gene has very limited methylation overall based on other studies we have done.
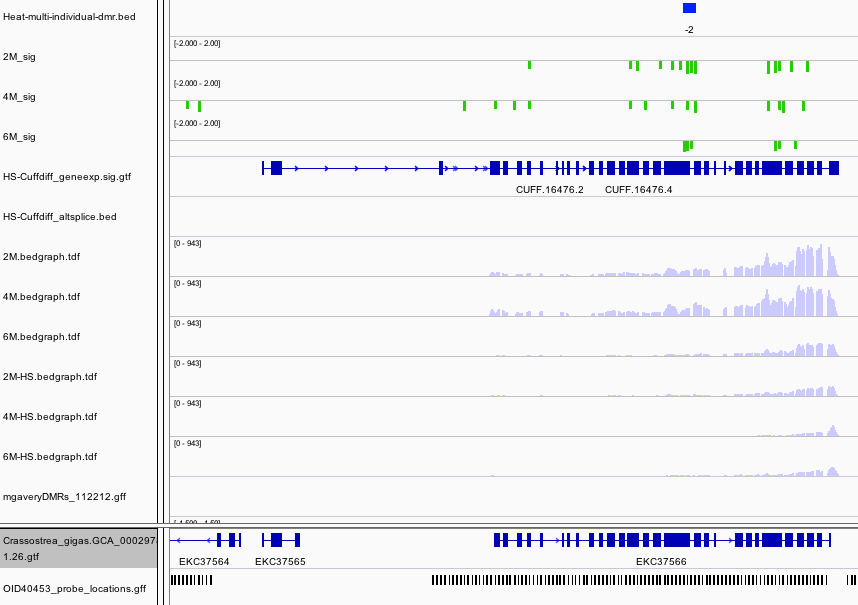
Another interesting DMR was found in Methylated-DNA–protein-cysteine methyltransferase.
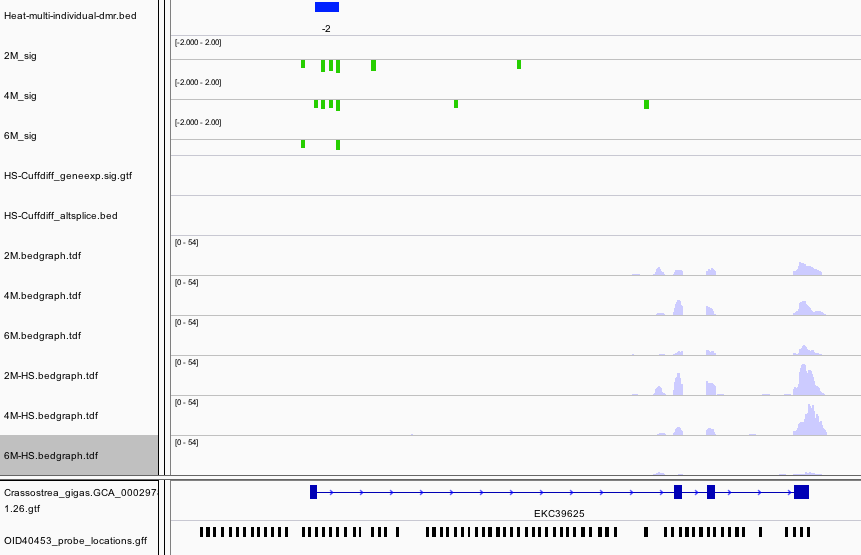
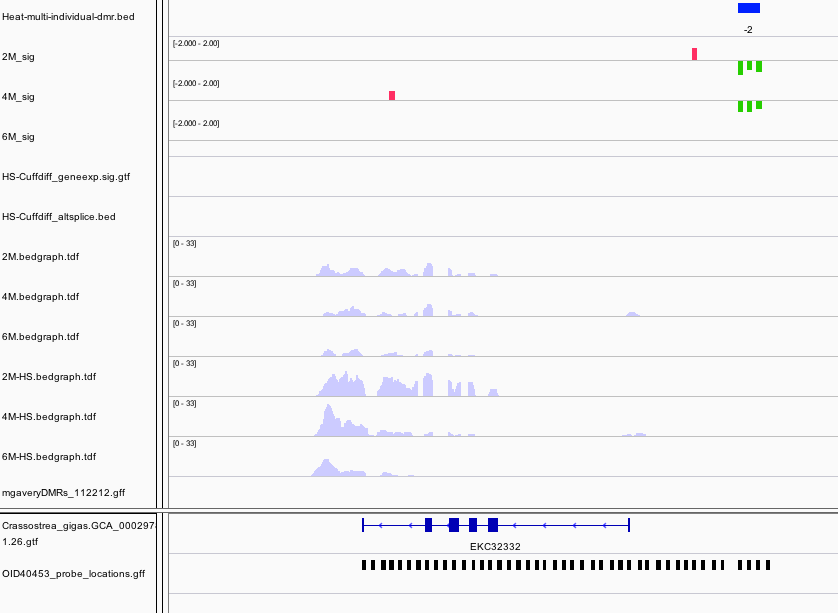
Within the intron of a Nacrein-like protein is a hypomethylated DMR. This is a negative regulator of calcification in shells of mollusks.
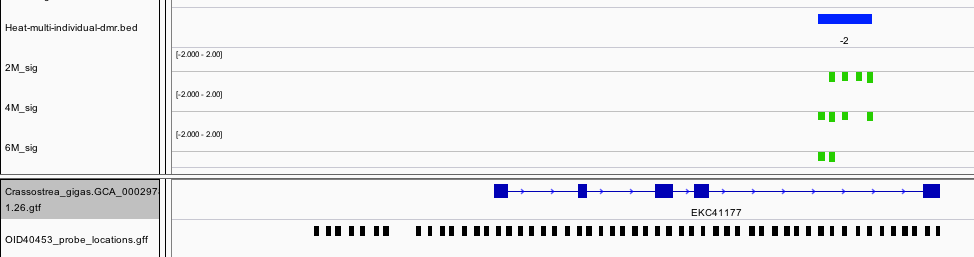
Collagen alpha-1(IV) chain is another gene that contains a hypomethylated DMR. This protein is the major structural component of basement membranes.
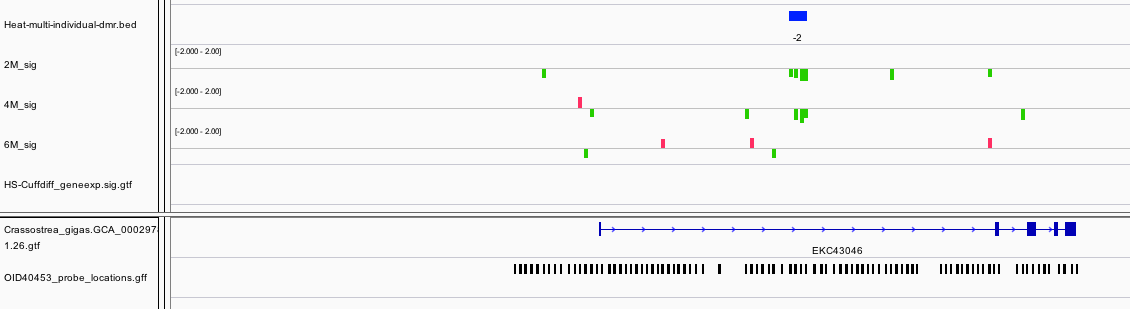
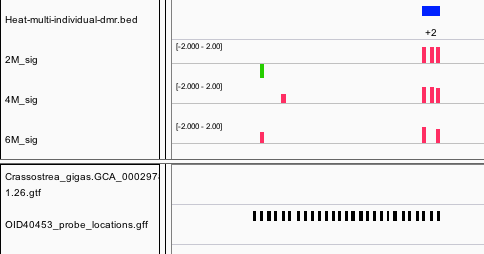
The only DMR that is hypermethylated is odd in the fact that annotation was dropped one the data was integrated into Ensembl. This could be related to the fact that the closest blast hit to this gene model is Insertion element IS1 protein insA and transposable element in prokaryotes.















Recent Comments